研究内容ー神経発生
脳神経系の発生過程では、比較的均一な神経幹細胞から極めて多様性に富んだニューロンやグリア細胞が分化します。この神経分化過程は転写因子によって厳密に制御されると考えられていますが、哺乳動物では詳細はよくわかっていません。本研究室ではこの神経分化過程を次の4つのステップに分けて、それぞれを制御する転写因子を同定し、神経分化における遺伝子発現制御ネットワークの解明を目指しています。
・神経幹細胞の増殖・維持
・神経幹細胞からニューロンへの分化
・神経幹細胞からグリア細胞への分化
・ニューロンのサブタイプの決定
方法としては、レトロウイルスベクターやエレクトロポレーション法を用いたgain of function解析とノックアウトマウスを用いたloss of function解析を行っています。これらの解析から上記の4つのステップを制御する転写因子が徐々に明らかになってきました。
最近の解析から、神経幹細胞の維持、ニューロンの分化、グリア細胞の分化のいずれのステップもbHLH因子(basic region-helix-loop-helix構造を持つ転写制御因子)が重要な役割を担うことが明らかになってきました(図1)。
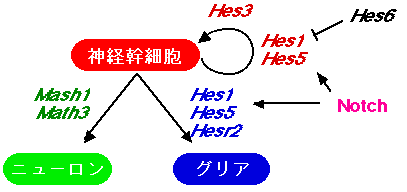
図1:神経分化を制御するbHLH因子ネットワーク
神経幹細胞の維持、ニューロンの分化、グリア細胞の分化のいずれのステップもbHLH因子が重要な役割を担っています。
1. 神経幹細胞の増殖・維持
bHLH因子Hes1やHes5を強制発現させると未分化状態を維持させることができ、逆に両遺伝子を欠損させると未分化状態の維持が阻害され、正常よりも早くニューロンへの分化がおこります。すなわち、Hes1とHes5は神経幹細胞の維持に必須の役割を持っています(Ohtsuka et al., 1999; 2001; Takatsuka et al., 2004; Hatakeyama et al., 2004)。また、Hes3にも同様の活性があります(Hirata et al., 2000; 2001)。Hesによく似たbHLH因子Hesr1, Hesr2も神経幹細胞を維持する活性があることがわかりました(Sakamoto et al., 2003)。
2. 神経幹細胞からニューロンへの分化
bHLH因子Mash1やMath3を強制発現させると神経幹細胞からニューロンへの分化がおこり、逆に両遺伝子を欠損させるとニューロンへの分化がおこりません。すなわち、Mash1とMath3はニューロンの分化決定因子として機能します。また、両遺伝子が欠損すると、本来、ニューロンに分化するべき細胞は幹細胞の状態でずっと止まるのではなく、グリアに運命転換することがわかりました。したがって、Mash1とMath3はニューロンの分化を決定するだけでなく、グリアの分化を積極的に抑制します(Tomita
et al., 2000; Ohsawa et al., 2005)。
また、Hes6はHes1とよく似た構造をしていますが、Hes1の機能を抑制し、Mash1がHes1に打ち勝ってニューロンに分化させるのを助けます(Bae
et al., 2000)。一方、Hesによく似たbHLH因子HeslikeはMash1と協調してGABA産生ニューロンの形成を促進します(Miyoshi et al., 2004)。
3. 神経幹細胞からグリア細胞への分化
bHLH因子Hes1やHes5は発生の初期には神経幹細胞の維持に作用しますが、発生の後期にはグリアへの分化を促進します(Hojo et al., 2000)。また、Hesとよく似た構造をもつbHLH因子Hesr2もグリアへの分化を促進します(Satow et al., 2001)。
4.ニューロンのサブタイプの決定
bHLH因子Mash1やMath3はニューロンの分化を促進しますが、サブタイプまで決めることはできません。サブタイプの決定にはbHLH因子とホメオボックス因子の組み合わせが重要です(Hatakeyama et al., 2001・Inoue etal.,2002)。
当研究室のテーマ
 English
English